Principles of ecology
Commuity ecology introduction
Principles of Ecology
Logistics reminders
- Semester project potential formats due at the end of next week
- In-class activity this Friday
- Grading in Moodle: Reflection 1 done; Gaurav will (re)do In-class activity 1 this week.
So far…
Dynamics of population growth
The simplest model (exponential growth) did not account for individuals interacting with one another (constant \(b\) and \(d\))
The logistic growth model accounts for negative interactions between individuals, which causes \(d\) to increase and/or \(b\) to decrease as populations get bigger.
- This gives rise to a maximum stable population size (\(K\), or Carrying capacity)
Logistic model with Allee effects account for positive and negative interactions. These case growth rate to increase with population size when populations are small, but decline when populations get big.
- This gives rise to a “minimum” or “threshold” population size, below which the population is expected to go extinct.
Next unit
Community ecology adds further complexity: individuals of the same species can interact, but individuals of different species can also interact.
This can get complicated:
- Lots of species in any given community
- Lots of different types of interactions
- Interactions can change in different contexts

“The organism as the subject and object of evolution”
- “Organisms are both the consumers and the producers of the resources necessary to their own continued existence.”
White pine trees in New England make such a dense shade that their own seedlings cannot grow up under them, so hardwoods come in and take their place.
On the other hand, organisms may make an environment more hospitable to themselves.
Grazing animals actually increase the rate of production of forage, both by fertilizing the ground with their droppings, and by stimulating plant growth by cropping.
Beavers create ponds by felling trees and building dams; indeed, a significant part of the landscape in northeastern United States has been created by beavers.
The activity of any organism changes the environment in which it lives.
- The movement of an earthworm through soil aerates the soil
- The transpiration of a tree cools the atmosphere
- An organism may add or subtract resources that are available to other organisms
- In some cases, an individual will actually enter into the lives of other organisms
This can in turn shape the dynamics of other species.


Community ecology
- The study of how interactions between species shape ecological communities
- Vast subject, given the wide variety of interactions that can unfold in nature
Keep track of the interactions in this video, and consider their consequences
Direct and Indirect interactions
For an hourlong documentary about hummingbirds
https://www.youtube.com/watch?v=yc8oRjt7jpkDirect and Indirect interactions

Direct and Indirect interactions
Direct and Indirect interactions

Conceptual model of a pollination network from MacGregor et al. (2023), Ecological Entomology
Direct and Indirect interactions
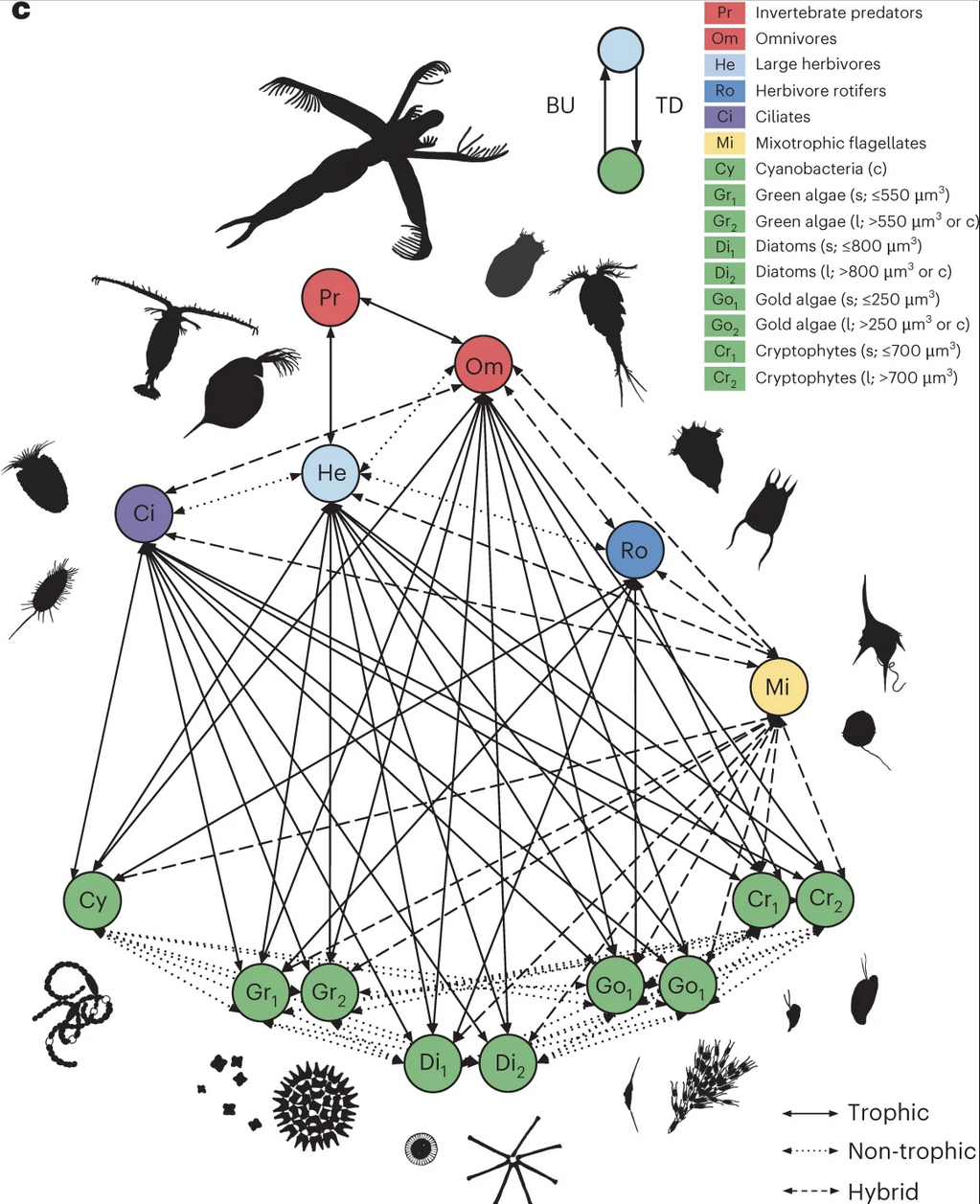
Conceptual model of a plankton network in temperate from Merz et al. (2023), Nature Climate Change
Approaches to community ecology
- Reductionism
- Study a complex system by breaking it down into component interactions
Photo from Smithsonian’s Tropical Research Institute
e.g. In a forest that houses hundreds of tree species, how do two interact with each other?
Approaches to community ecology
- Reductionism
- Study a complex system by breaking it down into component interactions
- e.g. In a forest that houses hundreds of tree species, how do two interact with each other?
- Where else do biologists take a reductionist approach?
Approaches to community ecology
- Holism
- Historically, some ecologists have favored a view of communities as “superorgansims”
- “Emergent properties” that are only evident when we study complex, messy networks
- Where else do biologists take a holistic approach?
Approaches to community ecology
In this class, we will explore both approaches…
- Reductionist approaches, e.g.
- Under what conditions can a pair of species that competes with one another for the same resources coexist?
- How do predator–prey interactions shape population dynamics?
- Under what conditions can a pair of species that competes with one another for the same resources coexist?
- Holistic approaches, e.g.
- How do disturbances (re)shape the structure of ecological communities?
- Under what conditions do interactions between species change from competitive to mutualistic?
Community ecology, day 2
Approaches to community ecology
Last time in class, we discussed reductionist and holistic approaches to wrestling with the complexity of ecological communities.
- We will begin with the reductionist approach in this class, which has been a longstanding tradition across subdisciplines of biology.
Reductionist approach:
Ecological communities as composites of pairwise interactions
- What are the ways in which two species can interact?
Nature of species interactions
| Sp 1 effect on Sp 2 |
Sp 2 effect on Sp 1 |
Shorthand |
|---|---|---|
| Benefit (+) | Benefit (+) | |
| Harm (–) | Harm (–) | |
| Benefit (+) | Harm (–) |
Nature of species interactions
| Sp 1 effect on Sp 2 |
Sp 2 effect on Sp 1 |
Shorthand |
|---|---|---|
| Benefit (+) | Benefit (+) | Mutualism |
| Harm (–) | Harm (–) | Competition |
| Benefit (+) | Harm (–) | Predation |
Nature of species interactions
| Sp 1 effect on Sp 2 |
Sp 2 effect on Sp 1 |
Shorthand |
|---|---|---|
| Benefit (+) | Benefit (+) | Mutualism |
| Harm (–) | Harm (–) | Competition |
| Benefit (+) | Harm (–) | Predation Herbivory Parasitism |
Nature of species interactions
| Sp 1 effect on Sp 2 |
Sp 2 effect on Sp 1 |
Shorthand |
|---|---|---|
| Benefit (+) | Benefit (+) | Mutualism |
| Harm (–) | Harm (–) | Competition |
| Benefit (+) | Harm (–) | Predation Herbivory Parasitism |
| Neutral (0) | Benefit (+) | Commensalism |
| Neutral (0) | Harm (–) | Ammensalism |
To further complicate matters, the same pair of species can have different interactions under different conditions
Recall the harvest mouse from previous class
- Let’s sketch a network of interactions
Your job (individually): Sketch a similar network for one of the communities you are considering working with.
Who are the major players?
How are they related to one another?
What are some ways that human activity might perturb the network?
Group discussion
Please nominate within each group:
Facilitator: Manage group discussion time (~5 minutes per person); ensure that everyone gets a chance to speak/ask questions; encourage conversation if things get quiet
Recorder: Document the conversations and share notes with relevant individuals (e.g. if we are discussing my idea, Recorder would keep track of the questions that come up and share that with me after the group discussion)
Reporter: Share group’s ideas with the class - what are some of the general themes across the group, what challenges do you as a group or individuals envision, etc.
Within groups, discuss…
What idea(s) you had going into this assignment, and why?
From your literature search, do you feel that your idea was too narrow/broad/just-right/other?
Among the primary literature you are finding, is there a good mix of “basic” ecology and more “applied” environmental or social science?
From your searches so far, do you see ways to connect material from class to your semester project? (either material so far, or from future topics)
What paper/idea/resource/learning surprised you the most?
What do you think are your next steps for either (a) selecting a focal community from among your options, (b) or getting more information on the community you have decided to focus on?
Community ecology, day 3
Population ecology review
In the population ecology modules, we explored three basic model structures
Exponential growth: \(\frac{dN}{dt} = rN\)
- Stage-structured populations also grow exponentially, but the rate of growth \(r\) is more complicated
Logistic growth: \(\frac{dN}{dt} = rN(1-\frac{N}{K})\)
- No matter initial population size, populations reach carrying capacity and equilibrate there
Logistic growth with Allee effects: \(\frac{dN}{dt} = - rN \bigg( 1-\frac{N}{T} \bigg) \bigg( 1-\frac{N}{K} \bigg)\)
- If populations stay above threshold \(T\), they grow to carrying capacity
- But if they fall below \(T\), they decline to extinction
We discussed the stability of these equlibrium points
Incorporating species interactions
- We will begin with the logistic growth model as our “baseline”:
\[\frac{dN_1}{dt} = rN_1(1-\frac{N_1}{K_1})\]
- Alternative expression:
- If we set \(\alpha_{11} = \frac{1}{K_1}\), we can rewrite the logistic growth equation:
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1})\]
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1})\]
\(\alpha_{11}\) is the strength of within-population competition (also called intraspecific competition)
When there is competition between species, the growth rate of species 1 (\(\frac{dN_1}{dt}\)) is also affected by species 2
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1} - \text{competition from species 2})\]
\[\frac{dN_2}{dt} = rN_2(1-\alpha_{22}{N_2} - \text{competition from species 1})\]
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1} - \text{competition from species 2})\]
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1} - \alpha_{12}N_2)\]
\[\frac{dN_1}{dt} = rN_1(1-\alpha_{11}{N_1} - \alpha_{21}N_1)\]
We will focus on this set of equations next week
- What is the equlibrium for this system?
- When can species coexist, and when would one be expected to outcompete the other?
- Even with this “reductionist” approach… things get tricky!